
Wednesday, February 14, 2007
Tuesday, November 21, 2006

Thursday, July 20, 2006
You Know
I've found a very informative website, which deserves mention in both my sidebar and in its very own post. The site is called Nigga Know Technology, and is full of helpful reviews of consumer items and services. For example, a review of Garmin's new Portable FishFinder:
----
Apparently, when a white motherfucker go out in they boats they need to know where the fish at. This gadget fires out some of that radar and blows all that data up on the display. So those pasty niggas row they asses out where ever the fuck Garmin tells them to row they asses to and they throw they lines in. Must be a motherfucking blast for white people, but for serious that sounds like some boring ass shit...
asses out where ever the fuck Garmin tells them to row they asses to and they throw they lines in. Must be a motherfucking blast for white people, but for serious that sounds like some boring ass shit...
*BLIP*
*BLEEP*
THE FISH TO THE RIGHT MOTHERFUCKER
To sum this shit up, we don't know a fish finder because, niggas aint trying to find no fish. We leave that shit to the sleepy heads and those country ass Huckleberry Finn niggas. If Garmin wanna boost they nigga profits on they line of Finder units then Garmin gotta help a nigga find shit a nigga looking for like white breezies or the identity of that nigga who pretending to be TUPAC on them new cuts. Any way I'm out on that shit.
----
Other reviews focus on remote controls, 'jacks', and downloadable movies. All very helpful, and worth your free time.
----
Apparently, when a white motherfucker go out in they boats they need to know where the fish at. This gadget fires out some of that radar and blows all that data up on the display. So those pasty niggas row they
 asses out where ever the fuck Garmin tells them to row they asses to and they throw they lines in. Must be a motherfucking blast for white people, but for serious that sounds like some boring ass shit...
asses out where ever the fuck Garmin tells them to row they asses to and they throw they lines in. Must be a motherfucking blast for white people, but for serious that sounds like some boring ass shit...*BLIP*
*BLEEP*
THE FISH TO THE RIGHT MOTHERFUCKER
To sum this shit up, we don't know a fish finder because, niggas aint trying to find no fish. We leave that shit to the sleepy heads and those country ass Huckleberry Finn niggas. If Garmin wanna boost they nigga profits on they line of Finder units then Garmin gotta help a nigga find shit a nigga looking for like white breezies or the identity of that nigga who pretending to be TUPAC on them new cuts. Any way I'm out on that shit.
----
Other reviews focus on remote controls, 'jacks', and downloadable movies. All very helpful, and worth your free time.
Sunday, July 09, 2006





Tuesday, May 09, 2006
Her Voice is Able

The third record by Jolie Holland was released to music stores today, so they can sell it to you and you can go "coo, coo". Jolie is a much-admired singer and songwriter originally from Texas, but then she moved to San Francisco, where she lives now with 400 puppies and snakes. She's true blue, if that's important to you. I once met her after a show in St. Augustine and told her she was amazing. She said, "Precious."
A couple mp3's are around her myspace page, as long as they're around.
Sunday, April 23, 2006

Monday, April 03, 2006
Head Check

The measurement of brain volumes is a wide and imprecise science that has something in common with the type of generalist thinking that engenders sciences like phrenology or palmistry. Which aren't sciences. Because of the broad correlations between brain size, complex behavior, and evolution, a number of analyses have been applied to investigate these assumptions underlying the development of intelligence and the emergence of cognition. We begin with the assumption that bodies are ‘run’ by their brains, and that an increase in body size would lead to an increase in size of the controlling organ. Also within the background of our thinking is the idea that bigger brains equal better brains: more neurons, more processing, more developed intelligence. But how can we represent these general ideas in a more careful analysis that tells us something significant about brains and behavior?
The devil, of course, is in the details, and it is likely the constituents characteristics of a nervous system that may reveal the significant volume-dependent factors for the emergence of cognition. Cortical folding, lobe development and more extensive brain-body scales have come to describe how the brain as an organ might have anything to do with the body. Fifty years ago Von Bonin evaluated the index of cortical folding (IFC) as a ratio of the total cortical surface (unfolded) to the exposed cortical surface. The measure, however, is largely confounded as a marker of ‘intelligence’ by generalizations across cladistic orders; as a group, cetaceans have evolved extraordinarily high IFCs, larger than most mammals, yet not all whales and dolphins are seen as of the same intelligence. In order to parse apart these differences within orders, the 19th century neuroanatomist Olaf Snell devised an equation using specific ‘scaling effects’ to find the encephalization quotient (EQ) of particular species,
E=csr
where E is the weight of the brain, S is the body weight, C is a constant ‘cephalization factor’, and r is an empirically determined exponential constant for a given order. The EQs for various mammalian species include:
Dolphins 5.31
Chimps 2.47
Rhesus monkeys 2.09
Elephant 1.87
Whale 1.76
Dog 1.17
Cat 1.00
Rat 0.40
Though crude, the equation does help represent a statistic that better reflects relative brain size and cognitive intelligence. Man ends up at the top of the heap with an EQ of 7.44, while dolphins and chimps follow after, supporting the large comparative literature that uses those subjects as models for ‘intelligent behavior’. I remain glibbly mum about what it says about cats.
A problem with the use of these measures, however, is that they all treat the brain as a unitary organ; cognitive neuroscience posits that component processes are driven by differential neural activity. In order to make assumptions about intelligence we must define what component processes (and component brain areas subsuming those processes) inform our notions of intelligence and awareness. Would an animal with a brain that is 20% of its body mass but 97% sensory & motor cortex be smarter than an animal with more primate-like proportions? Even regionally-based analyses of comparative neuroanatomy have their own problems. As regions in the PFC become more functionally defined (e.g., the DLPFC is a ‘working memory’ area), comparative distinctions become more difficult. Behavioral analogues of executive functions are notoriously difficult to find even in higher primates, let alone species that might share a similar IFC or EQ across cladistic orders.
And, of course, why are the measurements of relative or absolute brain sizes important, besides inflating our own species’ egos? Each species evolves a brain size to its need. In order to represent the broad taxonomic trends that follow the adaptations of these needs, measures of brain volume can inform us about the general trends of emergent cognition among species, but they say little about specific within-species variances. A biological statistician looking at a Jerison plot of brain mass/body mass (see the picture) might notice the conspicuous lack of error bars or SD units; given the variability in both brain size and body weight between members of the same species (males vs. females, zoo vs. wild animals, age differe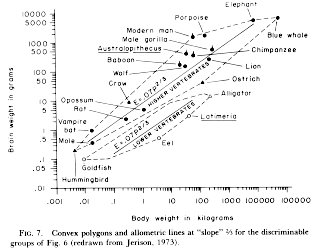 nces), it may be premature to make assumptions about the amount of variability within or across species. Another problem with relative brain size is the bias introduced in measuring it. For example, take the differences in relative brain size between bats whose diet consists mainly of fruit (frugivores) or leaves (folivores). Regardless of the effects of the diet on brain anatomy, the way each of these foodstuffs are digested affects the calculation of relative brain size: folivores have big stomachs and retain their food for longer in order to digest, thus increasing the total body weight and decreasing the relative brain size.
nces), it may be premature to make assumptions about the amount of variability within or across species. Another problem with relative brain size is the bias introduced in measuring it. For example, take the differences in relative brain size between bats whose diet consists mainly of fruit (frugivores) or leaves (folivores). Regardless of the effects of the diet on brain anatomy, the way each of these foodstuffs are digested affects the calculation of relative brain size: folivores have big stomachs and retain their food for longer in order to digest, thus increasing the total body weight and decreasing the relative brain size.
Where the two fuzzy measures come together is even harder to evaluate. Perhaps there is both some requisite amount of absolute brain mass and some specific cladistic orders that engender cognition. Perhaps we can make a preliminary guess that absolute brain size is best represented by environmental needs (or niches) and the relative brain size by the principles of Hebbian learning. Specifically, animals who meet the requisite absolute brain size may then gone evolve to involve larger brains relative to either body size and possibly a larger intelligence, all on a ‘use it or lose it’ basis, though we shouldn’t always assume that these measures are correlated.
Subscribe to:
Comments (Atom)
